18.2 전사개시의 조절 : 세균
- (DNA) lac operon 은 활성인자, 억제인자의 신호에 반응하여 발현을 조절한다.
1. lac 유전자의 조절 (lacZ, lacY, lacA)

- lac Z의 5‘에 위치한 lac 프로모터는 3개 유전자를 하나의 mRNA로 전사한다.
- 이 mRNA는 1개 이상의 유전자를 가지고 있기 때문에 다유전성 메시지라고 한다. (1개 mRNA에 3개 단백질 번역)
- lacZ는 lactose를 galactose와 glucose로 분해하는 효소를 암호화. 에너지원으로 이용
- lacY는 lactose를 세포안으로 운반하는 단백질을 암호화. 젖당투과효소
- lacA는 lactose 운반시 독성인 Thiogalactoside 제거
- lacZ, Y, Z 유전자는 lactose 있고, glucose 없을 때만 높은 수준으로 발현
- 이 과정에서 CAP (활성단백질)과 Lac 억제인자가 관여
- Lac 억제인자 : lacI 유전자에 의해 암호화. 다른 lacZ,Y,A 유전자 근처에 위치. 그러나 독립적으로 전사된다. lactose 없을 때만 DNA와 결합.
- CAP 단백질 : 분해활성 단백질. CRP (단백질수용체 cAMP) 로도 알려져 있다.
CAP 유전자는 lacZ,Y,A 와 인접해있지 않다. glucsoe 없을 때만 DNA 결합
- Lac 억제인자, CAP은 DNA 결합단백질. lac프로모터나 특정 DNA 부위에 결합
2. CAP 과 Lac 억제인자의 상반적 작용

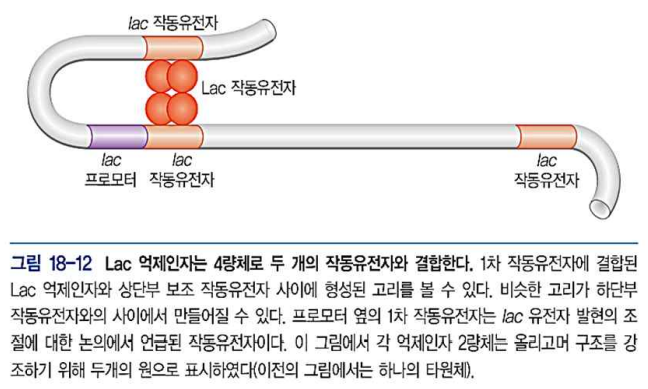
- Lac 작동유전자 (operator) : Lac 억제인자가 결합하는 DNA 부위. 두 소단위에 의해 인식됨. 21bp로 이루어진 이중대칭구조. Operator는 프로모터와 겹쳐있어서 억제인자가 RNA pol을 프로모터에 달라붙지 못하게 함.(CAP 이 없으면 억제인자 없어도 프로모터와 약하게 결합)
- lac 프로모터의 -35 서열이 최적서열 아니고, lac 프로모터에 활성증진부위(Up-element)도 없다.
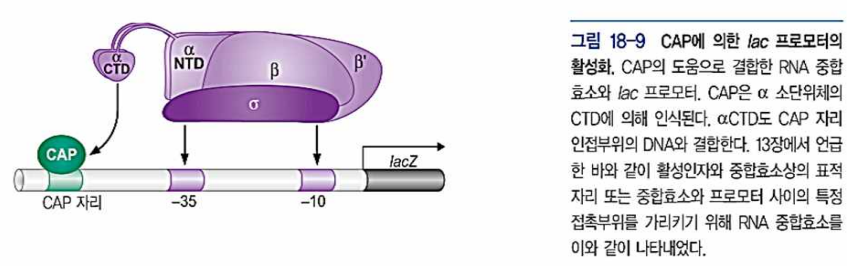
- CAP은 Operator와 길이가 비슷하지만 염기서열이 다르다. CAP은 다른 부위에 2량체로 결합. 60bp 위에 결합. RNA pol과도 결합하여 RNA pol을 프로모터로 끌어들임. 프로모터-RNA pol 결합 안정화
3. CAP의 구조적 특징
- CAP은 서로 분리된 활성표면(RNA pol의 α-CTD 와 결합) 과 DNA 결합표면을 가진다.
- 활성증진부위가 없는 lac프로모터에서 α-CTD는 CAP 단백질과 부근의 DNA에 결합.(DNA 큰홈이 인식하도록 도움을 줌)
4. CAP과 Lac 억제인자의 DNA 결합
- CAP과 Lac 억제인자는 공통구조모티프를 이용하여 DNA와 결합. DNA 큰홈을 인식.
- 일반적으로 단백질은 역반복이나 동형 2량체로서 결합 (2개 소단위)
- 단량체는 염기의 각 반쪽자리에 대칭으로 결합. 결합자리에 2량체로 있게 된다.
- 특정 DNA 서열 인식은 ⌜나선-회전-나선(HLH)⌟ 2차구조 모티프를 사용한다.
- 이 영역은 두 개 α나선으로 이루어져 하나는 인식나선으로 DNA 큰홈에 꼭 맞는다.
- α나선은 바깥면의 아미노산 잔기들이 염기쌍의 화학잔기들과 결합한다.
- 단백질은 이 방법으로 DNA 풀림 없이 DNA 서열을 구별할 수 있다. (큰홈)
- 나선-회전-나선의 2번째 나선은 DNA 골격과 접촉한다.
- 인식나선을 적절히 자리잡게하며 동시에 단백질-DNA 결합에 결합에너지를 제공한다.
(박테리오파지 λ 억제인자와 차이가 있다.)
- Lac 억제인자는 4량체로 결합하지만 각 Operator는 단 2개와 접촉
- 끼어있는 DNA는 고리를 형성하여 반응을 용이하게 한다.
- λ 억제인자는 아미노말단을 이용해 DNA와 추가접촉을 한다. (나선의 뒷면인 작은홈과도 결합)
- 일반적으로 단백질-DNA 결합이 DNA 구조를 변경시키지는 않는다.
- 일부의 경우, 단백질-DNA 복합체에서 DNA를 변형시킨다. (ex. CAP)
- CAP는 DNA를 크게 굽혀서 DNA가 단백질 둘레를 감싸도록 한다. (결합의 안정성 부여)
- 작동유전자 DNA의 꼬임을 일으키는 경우도 있다.
5. Lac 억제인자와 CAP 활성 조절
- Lac 억제인자, CAP 활성은 다른자리입체성으로 조절
- Lactose가 세포로 들어가면 Alloactose로 전환. (Lac 억제인자는 Lactose 말고 Allolactose로 조절)
- Lactose의 Allolactose 전환은 β-Galactosidase에 의해 촉매
- lac 유전자 발현이 억제되어도 약간씩은 발현된다. β-Galactosidase 를 조금씩 만든다
- 종종 RNA pol 이 Lac 억제인자 대신에 프로모터에 결합하기 때문이다.
- Lactose 가 없어도 세포에 낮은 수준의 젖당분해효소가 존재
- Allolactose는 Lac 억제인자와 결합하여 억제인자의 형태를 변화시킨다. → 억제인자는 DNA 결합 못하게 됨. lac 유전자 전사 시작!
- CAP 활성 : CAP 과 cAMP가 복합체를 만들면 DNA에 결합. glucose는 cAMP의 세포 내 농도를 낮춤. cAMP는 CAP의 다른자리입체성 효과인자이다. 포도당이 낮을 때 (cAMP 수준 높아짐) CAP은 DNA와 결합. lac 유전자 활성. 효과인자 cAMP와 결합하는 CAP 부위는 DNA 결합자리와 분리되어 있다.
6. 조합조절
- lac 유전자는 2개 신호에 의해 발현 (Lac 억제인자, CAP)
- E. coli 의 gal 유전자는 galatose 대사에 관여하는 효소를 암호화
- 기질 (=galactose) 존재, glucose 없을 때 발현
- CAP을 동일하게 활성인자로 사용 (조합조절, 통합조절). lac, gal 등 여러 유전자 조절
- 같은 신호가 여러 유전자들을 조절
7. 대체 σ 시그마 인자
- 프로모터 서열을 인지 : RNA pol의 σ 소단위체
- lac 프로모터는 σ70 소단위체를 가진 RNA pol 에 의해 인지
- 그러나 E.coli는 특정 환경에서 σ70 소단위체를 대체하여 다른 프로모터에 결합시키는 다른 σ소단위체를 만든다.
- 대체 σ인자 중 하나는 ⌜열충격 σ 인자⌟인 σ32 이다. (열충격받으면 σ70→σ32)

8. NtrC 와 MerR 전사활성인자
- 다른자리입체성 효과로 작용하는 활성인자 (활성화되지 않는 RNA pol을 프로모터로 이끈다)
- NtrC는 pol 의 구조적 변형 → 열린구조복합체
- MerR도 RNA pol-프로모터 복합체에 작용하지만, DNA에 작용하는 활성인자의 다른자리입체성 효과에 구조 변형
9. NtrC 특징 (전사활성인자)

- CAP 처럼 분리된 활성화 영역, DNA-결합영역을 가진다. 특정 신호가 있을 때만 DNA와 결합.
- 그 신호는 낮은 질소 수준 → DNA 결합영역이 드러남
- 활성화되면 NtrC는 glnA 유전자 자리에 2량체로 결합
- 2량체 간의 단백질-단백질 결합을 통해 협동적으로 4개 glnA에 결합 (한 번 결합하면 나머지 3개 glnA에 NtrC 쉽게 결합)
- NtrC 없으면 glnA를 전사하는 RNA pol 은 닫힌복합체
- 활성화되면 NtrC는 σ 인자와 직접 결합
- 이 결합을 위해 활성결합자리와 프로모터 사이의 DNA가 고리를 형성 → 상호 결합 용이
- NtrC 자체는 ATP분해효소활성이 있음 → RNA pol 형태적 변화 유도 (닫힌복합체→열린복합체)
- NtrC-결합자리와 프로모터 사이에 IHF결합자리가 있다. IHF는 DNA를 구부려 NtrC 활성을 증가시킨다.

10. MerR 전사활성인자 특징

- DNA를 비틀어 전사 활성화
- 수은 존재 시 MerR은 1개 DNA-결합자리에 결합하여 merT 프로모터 활성화
- MerR은 merT 프로모터의 -10과 -35 사이 서열에 결합
- MerR은 RNA pol 과 결합한 DNA 반대쪽에 결합해서 RNA pol은 MerR과 동시에 프로모터에 결합
- merT 프로모터 -10과 -35 자리 사이 거리는 길어서 σ인자에 의해 인식되는 두서열요소는 최적상태로 분리되지 못한다 → 수은 없을 때 MerR은 프로모터에 RNA pol이 결합하긴 해도 전사개시할 수 없는 형태를 만듦 → 기본 수준의 전사조차 일어나지 않음
- MerR이 수은에 결합하면 MerR 전사활성인자는 DNA 프로모터의 중심이 비틀어져 형태적 변화
→ -10과 -35자리를 σ프로모터에서 발현되는 것처럼 복원시킴 → 전사개시 (DNA 구조 변경)
참고문헌 : Watson. (2014). 왓슨 분자생물학(7판). (주)바이오사이언스출판
'공부 > 분자생물학' 카테고리의 다른 글
| [왓슨 분자생물학] 19장 진핵세포의 전사조절-1 (0) | 2024.01.04 |
|---|---|
| [왓슨 분자생물학] 18장 원핵생물의 전사조절-3 (1) | 2024.01.03 |
| [왓슨 분자생물학] 18장 원핵생물의 전사조절-1 (1) | 2023.12.23 |
| [왓슨 분자생물학] 17장 생명의 기원과 초기 진화 (1) | 2023.12.23 |
| [왓슨 분자생물학] 16장 유전암호 (1) | 2023.12.23 |



